![]() アサガオの系統関係
アサガオの系統関係
従来の植物の形態に基づく分類法の他に、最近ではある遺伝子を種間で比較して系統関係を解析することが広く行われるようになった。これは多くの遺伝子は一定の頻度で突然変異(塩基置換)を蓄積することを利用している。よく用いられる遺伝子としては葉緑体遺伝子のrbcLやmatK、種々の核遺伝子などがある。前者は塩基置換しにくいため比較的遠縁、後者は近縁の関係を解析するのに用いられている。またより近い、種内の関係を解析するにはゲノム全体の多型をスキャンする必要があり、この目的にはRAPD(Random Amplified Polymorphic DNA)やAFLP(Amplified Fragment Length Polymorphism)などが用いられている。
アサガオの属するIpomoea属(サツマイモ属)を細かく分けて、学名をPharbitis nilにする説とIpomoea nilとしてその中のアサガオ節(section)に分類する説があるが、最近では後者の分類による方が多いようである。
![]() Ipomoea属の分子系統樹
Ipomoea属の分子系統樹
このIpomoea属は全世界で約500種から構成され、この属には一年生草本から多年生草本、木本まで存在し、非常に多岐にわたっている。実際にアサガオ種内と近縁種およびサツマイモを含むIpomoea属の14種について葉緑体遺伝子のmatK塩基配列を決定し、これらをNeighbor-Joinning法で比較して系統関係を解析した。
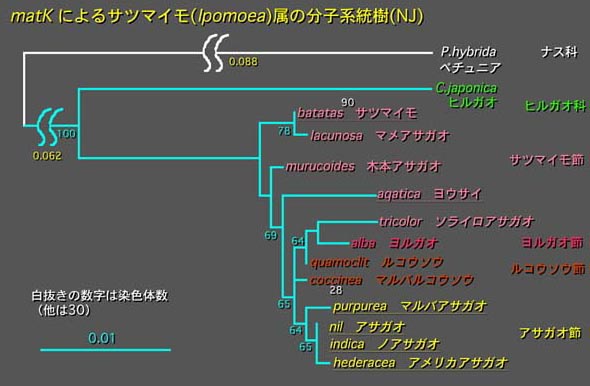
その結果、一年生草本、多年生草本、木本の種は明確に分かれた系統を作らなかった。このことは繁殖戦略の違いのような進化はそれほどドラスティックな変化ではなく、比較的少ないステップで変化しうるものかもしれない。Ipomoea属では一年性草本のほとんどは自家受粉するが、多年生草本はほとんどが自家不和合性であり、自分自身の花粉では受精できない。これはそれぞれの繁殖戦略によるものであることが容易に推測できる。すなわち、次年の種子を確実につくるためには自家受粉する必要があるが、多年生であればより多くの遺伝的変異を他の系統から導入するため自家不和合性であるほうが都合がよい。またアサガオはある程度の果実が結実すると枯死するようにプログラムされているが、例えば牡丹突然変異体は不稔であるため植物体の枯死が起こらない。このような過程を経て多年生となる進化が起こったのかも知れない。
![]() Pharbitis(アサガオ)節の分子系統樹
Pharbitis(アサガオ)節の分子系統樹
前述のmatK遺伝子による解析ではアサガオとその近縁種ではほとんど塩基置換がなかったため系統関係が不明であった。そのため、新規にクローニングしたC機能MADS-box遺伝子の一つである、牡丹(duplicated; dp)のイントロンを用いて分子系統樹を作成した。
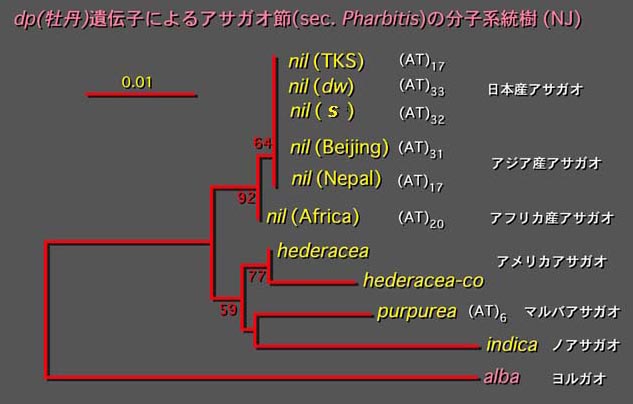
その結果、日本のアサガオおよび北京天壇・ネパール系などアジア産アサガオには内部のマイクロサテライト様領域のATの反復数は異なったが塩基置換はなかった。またアフリカ系は他のアサガオより他種に近く、祖先型であることを示している。このことはアフリカ系アサガオが他種と交配しやすいということとも一致している。
![]() AFLPによるPharbitis(アサガオ)節の分子系統樹
AFLPによるPharbitis(アサガオ)節の分子系統樹
より詳細にアジア産アサガオの系統関係を解析するため、ゲノム全体をスキャンすることで、より多くの多型を検出できるAFLP法(詳しくはマッピングの項目を参照)を用いて分子系統樹を作成した。
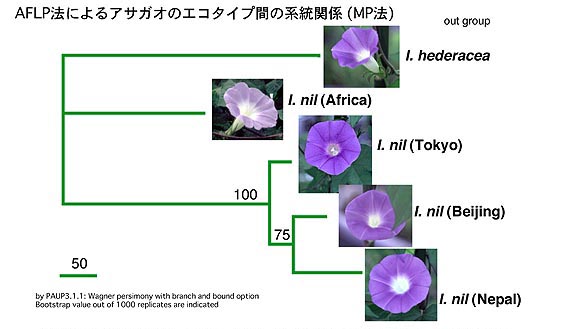
牡丹遺伝子を用いた系統樹と同様にアフリカ系がより他種(I. hederacea; アメリカアサガオ)に近いということが確かめられ、やはりこのことはアフリカ系がアサガオ(Ipomoea nil)の祖先種により近いことを意味している。
また日本産の東京古型標準型と一番近い系統は北京天壇系統であった。このことは奈良から平安時代にかけて中国から薬草としてアサガオが日本にもたらされたという事実と一致しており面白い。
よくアサガオの原種はどれか?とか、アサガオの起源はどこか?という論議があるが、種という意味ではこのアフリカ系なども立派なアサガオ(=Ipmoea nil)である。またアフリカ系とほぼ同一のブラジル系のアサガオが存在し、まだDNAは解析していないが、おそらく非常に近くなると思われる。中南米にIpomoea属の種数が多いことなどから考えあわせて、アサガオの起源は熱帯アメリカ地方であると考えられ、これがアジアに渡り、より鋭敏な短日感受性を獲得しながら温帯地方へと適当していったのではないだろうか。
 木になるアサガオ、Ipomoea
murucoides
木になるアサガオ、Ipomoea
murucoides{kind=link}